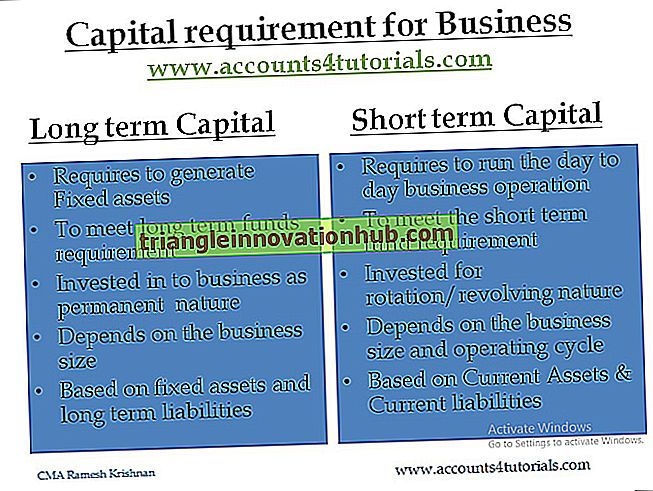
Struttura microscopica elettronica di una tipica cellula batterica
Struttura microscopica elettronica di una tipica cellula batterica!
Questi sono essenzialmente sistemi a una busta costituiti da materiale nucleare centrale circondato da citoplasma.
L'intera massa del protoplasto è racchiusa solo all'interno di una membrana plasmatica.
Le cellule batteriche mostrano una tipica struttura procariotica. È racchiuso da tre strati, il rivestimento più esterno della melma, la parete della cellula centrale e la membrana plasmatica più interna.
[I] Slime Layer, Capsule, Glycocalyx:
È uno strato gelatinoso presente sulla superficie esterna della parete cellulare, solitamente composto da polisaccaridi (es. Destrano, destrina, levan) e catene polipeptidiche di amminoacidi. Quando i costituenti di questo strato sono solo polisaccaridi che formano uno strato viscoso, esso viene definito come strato di melma. Ma quando le sostanze azotate (cioè gli amminoacidi) sono anche presenti insieme ai polisaccaridi, allora è chiamata capsula.

La produzione / formazione della capsula dipende da fattori ambientali e nutrizionali. Vari ceppi della stessa specie possono avere capsule di diversi polisaccaridi specifici del ceppo. Una diversa struttura, il glicocalice, è una massa aggrovigliata di sottili fibre polisaccaridiche che si estendono dalla superficie batterica. La capsula o glicocalice può svolgere diversi ruoli importanti.
1. Una capsula spessa può proteggere la cellula dalla disidratazione. Tuttavia, non impedisce il passaggio di nutrienti e prodotti di scarto solubili in acqua.
2. La capsula protegge alcuni agenti patogeni dall'ingorgo e dalla distruzione dei globuli bianchi del corpo, contribuendo così alla capacità di un organismo di causare la malattia.
3. A differenza della capsula, il glicocalice non è in grado di proteggere una singola cellula dalla distruzione. Tuttavia, può aiutare le cellule batteriche ad aderire l'una all'altra, formando un aggregato batterico che le cellule del sangue possono trovare difficile da inghiottire.
4. Il polisaccaride di glicocalice è responsabile di consentire ad alcuni batteri di attaccarsi alle superfici.
guaine:
Alcune specie di batteri, in particolare quelli provenienti da acque dolci e ambienti marini, sono racchiusi in una guaina, o tubulo. Le guaine sono composte da composti metallici insolubili, come ossidi ferrici e manganici, precipitati intorno alle cellule come prodotti della loro attività metabolica. La guaina non è una parte vitale della cellula.
gambi:
Alcune specie batteriche sono caratterizzate dalla formazione di un'appendice semi-rigida chiamata stelo che si estende dalla cellula. Consente alla cellula di legarsi a superfici solide con l'aiuto di una sostanza adesiva prodotta all'estremità distale.
[II] Parete cellulare:
La parete cellulare batterica è la struttura che circonda immediatamente la membrana cellulare. La parete cellulare è una struttura complessa composta da diverse sostanze. La rigidità della struttura della parete cellulare è dovuta principalmente al peptidoglicano (noto anche come mureina o muco-peptide), una sostanza presente solo nei procarioti.
Questi polimeri molto grandi sono composti da tre tipi di elementi costitutivi:
1. N-accetil glucosamina (NAG)
2. N-accetil muramic e (NAM) e
3. un peptide costituito da quattro o cinque amminoacidi, vale a dire, L-alanina, D-alanina, acido D-glutammico e lisina o acido diamminopimelico.
Gli zuccheri NAG e NAM si alternano per formare lunghe catene parallele. Attaccate alle molecole NAM sono catene laterali corte di quattro o cinque amminoacidi. Un singolo strato di peptidoglicano è una rete di catene zuccherine adiacenti legate insieme attraverso le catene laterali dell'amminoacido, creando così una struttura reticolata che copre l'intera cellula.
Alcuni batteri possono avere diversi strati di peptidoglicano. Gli antibiotici di penicillina e cefalosporina causano la morte alle cellule batteriche interferendo con la sintesi del peptidoglicano. I peptidoglicani, insieme ad altri due costituenti della parete cellulare, l'acido diamminopimelico e l'acido teichico, sono unici per i procarioti. È importante notare che gli amminoacidi nel peptide nei peptidoglicani esistono nella configurazione D.
I batteri Gram-positivi e Gram-negativi mostrano differenze nella composizione della parete cellulare. Le proprietà coloranti di questi batteri dipendono dalla loro composizione della parete cellulare. La resistenza agli antibiotici di alcuni ceppi dipende anche dalla composizione della parete cellulare.
[III] membrana citoplasmatica:
La parete cellulare è seguita dalla membrana plasmatica delle lipoproteine o dalla membrana citoplasmatica. Controlla il passaggio di sostanze chimiche in soluzione dentro e fuori la cellula. Fornisce inoltre i macchinari biochimici per spostare ioni minerali, zuccheri, amminoacidi, elettroni e altri metaboliti attraverso la membrana. La membrana citoplasmatica è un doppio strato di proteina fosfolipidica simile a quella presente nelle cellule vegetali e animali.
La principale differenza, tuttavia, è che non ci sono steroli nei procarioti. Gli enzimi respiratori (citocromi, enzimi del ciclo di Krebs, NADH, ecc.) Non sono legati separatamente dalle membrane presenti negli eucarioti: la superficie interna della membrana citoplasmatica stessa fornisce siti per l'attacco di questi enzimi.
La membrana cellulare è il sito di molte funzioni che includono:
1. Trasporto di molecole dentro e fuori la cellula
2. Secrezione di enzimi extracellulari,
3. Respirazione e fotosintesi,
4. Regolazione della riproduzione e
5. Sintesi della parete cellulare.
[IV] Citoplasma e inclusioni citoplasmatiche:
Il citoplasma batterico è una miscela complessa di carboidrati, proteine, lipidi, minerali, acido nucleico e acqua. Memorizza materiale organico sotto forma di glicogeno, volutina e poli-P-idrossibutirrato. Alcuni batteri fotosintetici e non fotosintetici accumulano anche zolfo e ferro nel loro citoplasma.
Le alghe blu-verdi sono in grado di utilizzare CO 2 e H 2 O, con l'energia fornita dal sole a causa della presenza di pigmenti fotosintetici che sono chiamati collettivamente "ficobiline", costituite da ficocianina (blu) e ficoeritrina (rosso) . I batteri fotosintetici come Rhodospirillum sp. e Chromatium sp. devono la loro abitudine autotrofa a un tipo speciale di clorofilla, batterioflorofilla.
Oltre alla porzione fluida e alle particelle di stoccaggio, il citoplasma batterico contiene anche un'area cromatica o nucleare e alcune altre inclusioni. La cellula batterica è priva di mitocondri, reticolo endoplasmatico, corpi centrosoma e Golgi.
1. Materiale nucleare:
La caratteristica di un nucleo batterico è l'assenza di membrana nucleare, nucleolo, chromonemata e linfa nucleare. Tale nucleo è chiamato nucleoide o genoforo. Sotto il microscopio elettronico il nucleoide sembra essere fibrillare e composto da un DNA a doppio o singolo filamento. Ha approssimativamente 5 x l0 6 coppie di basi e un peso molecolare di circa 3 x l0 9 .
La molecola del DNA è lunga circa 1000 μm, di solito forma una struttura ad anello o qualche volta rimane diffusa in tutto il citoplasma della cellula. Il nucloide di Escherichia coli ha un nucleo centrale di RNA, circondato da 12-82 supercoiling del DNA. Alcune molecole proteiche sono anche associate al DNA.
Il DNA batterico è privo di istoni quindi non può essere confrontato con i cromosomi delle cellule eucariotiche.
2. Plasmidi:
Le cellule batteriche contengono anche alcuni determinanti ereditari extra cromosomici che sono indipendenti dai cromosomi batterici o sono integrati in esso. Lederberg (1952) ha coniato il termine plasmidi per tali determinanti ereditari extra-cromosomici. I plasmidi portano solo geni non essenziali e non hanno alcun ruolo nella vitalità e nella crescita dei batteri. Quindi sono anche definiti come elementi autonomi superflui.
I plasmidi sono molecole di DNA circolare a doppio filamento e chiuse che variano in dimensioni da 1 Kb (basi Kilobase-1000) a maggiori di 200 Kb. Spesso i geni trasmessi da plasmidi codificano per gli enzimi che, in determinate circostanze, sono vantaggiosi per l'ospite batterico.
Tra i fenotipi conferiti da diversi plasmidi ci sono:
un. resistenza agli antibiotici
b. produzione di antibiotici
c. degradazione di composti organici complessi
d. produzione di coliche
e. produzione di enterotossine.
f. produzione di enzimi di restrizione e modifica
In condizioni naturali, molti plasmidi vengono trasmessi a nuovi ospiti mediante un processo simile alla coniugazione. Per la maggior parte, la replicazione del DNA plasmidico viene eseguita dallo stesso insieme di enzimi utilizzati per duplicare il cromosoma batterico. Alcuni plasmidi sono sotto "controllo rigoroso" il che significa che la loro replica è accoppiata a quella dell'ospite in modo che solo una o al massimo alcune copie del plasmide siano presenti in ogni cellula batterica.
D'altra parte, i plasmidi sotto "controllo rilassato" hanno numeri di copie di 10-200 che possono essere aumentati a diverse migliaia per cellula se la sintesi proteica dell'ospite viene interrotta. In assenza di sintesi proteica, la replicazione del plasmide rilassato continua mentre la replicazione del DNA cromosomico e dei plasmidi "stringenti" cessa.
3. Ribosomi:
Sparsi nel citoplasma sono numerosi ribosomi composti da RNA. Queste particelle sono siti per la sintesi proteica e si trovano in gruppi chiamati "polribosomi" o "polisomi" e sono costituiti da subunità grandi e piccole. I ribosomi batterici sono di 70 tipi S e consistono in due subunità.
La costante di sedimentazione della subunità più grande è 50 S e quella dell'unità più piccola è di 30 S. I primi hanno due molecole di RNA e 35 amminoacidi e l'ultima molecola di RNA e 21 diversi amminoacidi.
4. Mesosomi:
Questi sono complessi racconti localizzati della membrana citoplasmatica. Ci possono essere 2-4 mesosomi in una cellula e il numero è solitamente più alto nei batteri che mostrano attività respiratoria elevata, come i batteri nitrificanti.
Si trovano spesso a originarsi in un punto in cui la membrana inizia l'invaginazione prima della divisione cellulare e si attaccano alla regione "nucleare". Si ritiene che i mesosomi funzionino nella sintesi della parete cellulare e nella divisione del materiale nucleare.
[V] Flagella:
Il flagello batterico è una lunga appendice protoplasmatica a forma di frusta filamentosa che si sviluppa nell'involucro cellulare e si estende oltre la superficie cellulare. La struttura e la composizione dei flagelli batterici è completamente diversa da quella del flagello eucariotico. Invece della disposizione "9 + 2" di tubulina contenente microtubuli, c'è semplicemente un singolo filamento di proteina globulare chiamato flagellina.
È una struttura proteica nuda priva di microtubuli. Flagella è responsabile della locomozione delle cellule batteriche e è coinvolta nella chemiotassi e lavora come organo di senso di un batterio. Il flagello ha una lunghezza compresa tra 10 μm e 20 μm e ha uno spessore di circa 0.2 μm. Un flagello ha tre parti.
(a) Corpo basale o granulo basale
(b) Hook come struttura e
(c) Filamento
Non tutti i batteri possiedono flagelli come nei batteri atrichous (es. Pasteurella, Lactobacillus). I flagelli batterici sono rigidi e non si muovono avanti e indietro. Invece, girano molto come il propellar di una barca.
[VI] Pili (Singular-pilus):
Conosciuto anche come fimbriae (singolare-fimbria), questi possono essere visti solo in una micrografia elettronica. Sono più numerosi dei flagelli. Queste sono appendici filamentose piccole, dritte e sottili non associate alla motilità dell'organismo. Consiste di una proteina chiamata pilina sistemata elicoidalmente attorno a un nucleo centrale. Le loro funzioni conosciute includono:
1. Il tipo speciale di pilus chiamato F. pilus (pilus del sesso) funge da porto di entrata del materiale genetico durante l'accoppiamento batterico (coniugazione)
2. Servire come mezzo di attaccamento a varie superfici, cioè contribuire alla creazione di infezione legando i batteri alle superfici cellulari di animali e piante dai quali possono ottenere sostanze nutritive.
micoplasmi:
I micoplasmi sono i più piccoli procarioti aerobici noti senza una parete cellulare scoperta da Pasteur (1843) mentre stava studiando l'agente eziologico della pleuropolmonite nei bovini. Sono stati designati come PPLO (organismi simili alla pleuropolmonite). Nel 1898, Nocard e Roux, riuscirono a ottenere colture pure di questi microrganismi in terreni contenenti siero.
I micoplasmi sono frequenti contaminanti nelle colture di tessuti ricchi di sostanza organica. Sono stati trovati anche nelle sorgenti di acqua calda. Si verificano nel suolo, nelle acque di scarico e in diversi substrati e nell'uomo, negli animali e nelle piante.
Questi microrganismi pleuromorfi sono stati nominati ascoococcus mycoides di Borrel et al. (1910). Nowak (1929) collocò questo organismo nel genere Mycoplasma, che appartiene alla classe Mollicutes dell'ordine Mycoplasmatales. Il micoplasma può essere coltivato su terreni liberi dalle cellule.
Struttura dei micoplasmi:
L'assenza di una vera parete cellulare rende questi organismi altamente plastici e facilmente deformabili, quindi i micoplasmi sono irregolari e di forma variabile. Le cellule possono essere coccoidi, granulari, a forma di pera, a grappolo, ad anello o filamentose.
Questi organismi sono ricoperti da una membrana citoplasmatica di lipo-proteina unitaria, 7, 5-10 μm di spessore. Il citoplasma contiene ribosomi e struttura simile a nucleoplasma. Sebbene il materiale genetico sia composto sia da DNA che da RNA, è meno della metà di solito presente in altri procarioti ed è forse il limite più basso richiesto per l'organismo cellulare. La quantità di DNA è fino al 4% e l'RNA circa l'8%. Il contenuto di G + C nel DNA oscilla tra il 23 e il 40 per cento.

Le cellule PPLO hanno diversi enzimi utili nella replicazione del DNA, la trascrizione e la traduzione coinvolte nella sintesi proteica e la generazione di ATP dallo zucchero anaerobicamente.