Cromosomi procariotici: note utili sui cromosomi procariotici
Ecco le tue note sui cromosomi procariotici!
L'informazione genetica di una cellula procariotica viene trasportata nel nucleoplasma sulla struttura chiamata cromosoma batterico. Consiste in una doppia molecola di DNA elicoidale (acido desossiribonucleico), mai associata a proteine basiche, ed è stata dimostrata in alcuni procarioti come circolare.

Cortesia dell'immagine: hangthebankers.com/wp-content/chromosome.jpg
Il cromosoma batterico non è quindi strutturalmente omologo con i cromosomi nucleari della cellula eucariotica, ma piuttosto con il DNA organellare presente nei mitocondri e nei cloroplasti eucariotici. È probabile che un singolo cromosoma batterico (cioè una molecola di DNA molto lunga) porti tutte le informazioni genetiche necessarie a specificare le proprietà essenziali della cellula procariota.
I plasmidi: 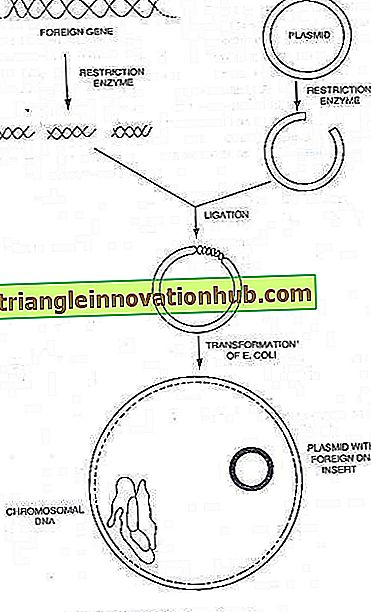

Molti batteri possono anche ospitare piccole molecole di DNA circolare cromosomiche in grado di replicazione autonoma, che sono conosciute come plasmidi finora investigate e portano i determinanti per tali caratteri fenotipici come resistenza ai farmaci e altre sostanze antibatteriche, e per gli enzimi che mediano determinate vie metaboliche . La quantità di DNA in un plasmide è compresa tra 0, 1 e 5 percento di quella nel cromosoma batterico.
È stato scoperto in Escherichia coli che ci sono due tipi di accoppiamento e durante la coniugazione un partner agisce solo come donatore genetico, o maschio, e l'altro solo come ricevente genetica o femmina. Poiché l'unica funzione del maschio è quella di trasferire parte del suo DNA, non è necessario che rimanga vitale, mentre la cellula femminile deve rimanere vitale, così che lo zigote possa svilupparsi in esso.
È stato scoperto che la virilità nei batteri è determinata da un elemento genetico trasmissibile quando i batteri maschili e femminili si coniugano, ogni cellula femminile viene convertita in un maschio. L'elemento genetico che governa la proprietà ereditaria della mascolinità è chiamato fattore F (F = fertilità), è trasmesso solo attraverso il contatto diretto tra cellule. Nel 1952 Lederberg coniò il termine plasmide come nome genetico per tutti i determinanti ereditari cromosomici supplementari, di cui F è un esempio.
È ormai noto che i plasmidi batterici sono piccole molecole circolari di DNA che trasportano i geni per la loro replicazione. In molti casi, portano anche geni che conferiscono nuove proprietà alla cellula ospite, come la resistenza ai farmaci o la produzione di tossine. Molti plasmidi portano geni che governano il processo di coniugazione. Quindi, la coniugazione è un meccanismo imposto alla cellula batterica da un plasmide, il cui risultato normale è il trasferimento del DNA plasmidico.
Tipi di plasmidi:
La maggior parte dei plasmidi sono stati classificati sulla base delle proprietà dell'ospite. Quindi, ci sono il fattore R (R = resistenza) e i fattori COI (COI = colicinogeny) dei batteri gram-negativi, i plasmidi penicillinasi di Staphylococcus aureus, i plasmidi degradativi di Pseudomonas, i plasmidi criptici ecc.
Proprietà dei plasmidi:
La struttura di tutti i plasmidi noti consiste di molecole di DNA a doppio filamento circolari. Molti di loro hanno pesi molecolari compresi tra 5 × I0 7 e 7 × 10 7 . Uno (un fattore R) ha un peso molecolare di solo 1 × 10 7, e alcuni dei plasmidi criptici sono ancora più piccoli. Poiché la quantità di DNA richiesta per codificare per un polipeptide medio con un peso molecolare di 40.000 è circa 6 × 10 7, F 1 e altri plasmidi di dimensioni simili possono contenere fino a 100 geni.
Espressione genica nei procarioti:
Cromosomi batterici:
È stato stabilito ora che un cromosoma batterico è sempre presente nel centro della cellula batterica che non è capace di divisioni mitotiche e meiotiche e non è racchiuso in una definita membrana nucleare. Una cellula batterica media contiene un millesimo del contenuto di DNA di una tipica cellula eucariotica.
Il cromosoma batterico è un sottile e allungato filamento flessibile e circolare della molecola di DNA associato a poche proteine. Le cellule batteriche possono dividersi molto rapidamente. La divisione cellulare è completata dal raddoppio di tutti i costituenti cellulari seguiti dalla partizione della cellula in due cellule figlie. Il primo passo nella divisione cellulare è la duplicazione del DNA.
Ricombinazione genetica nei batteri:
Questo è un processo in cui i materiali genetici, contenuti in due genomi separati, sono riuniti all'interno di un'unità. Nei batteri la ricombinazione avviene mediante (1) trasformazione, (2) trasduzione e (3) coniugazione. Questi fenomeni sono già stati discussi in dettaglio nei paragrafi precedenti, sotto il titolo "riproduzione sessuale nei batteri".

Induzione e Repressione:
Il potenziale genetico di un virus è limitato e utilizza essenzialmente il meccanismo host per la sua replica. D'altra parte, i batteri possono sintetizzare i loro costituenti da semplici sali e zuccheri. Il batterio, Escherichia coli ha circa 2500 geni sul suo cromosoma. Il batterio mantiene la sua economia interna solo permettendo l'azione di pochi geni alla volta.
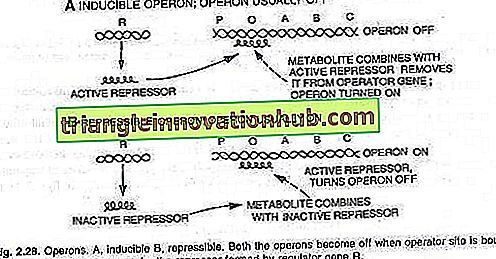
Ciò significa che i geni verranno attivati o disattivati secondo i requisiti. Una serie di geni verrà attivata quando è necessario gestire e metabolizzare un nuovo substrato. Quando viene attivato l'insieme di questi geni, vengono prodotti enzimi che metabolizzano il nuovo substrato. Questo fenomeno è chiamato induzione e le molecole (metaboliti) che evocano questo fenomeno sono induttori.
D'altra parte, quando un metabolita necessario al batterio viene fornito in eccesso dall'esterno, il batterio inibisce la sua produzione e questo impedisce l'ulteriore produzione del metabolita da parte del batterio. Questi geni inattivati sono quindi denominati repressibili e il fenomeno è noto come repressione o repressione del feed-back.
Tuttavia, alcuni dei geni sono costantemente espressi per prendersi cura della normale attività cellulare come la glicolisi. Questi geni sono noti come costitutivi e gli enzimi prodotti da loro sono noti come enzimi costitutivi, come la deidrogenasi.
Concetto di Operon:
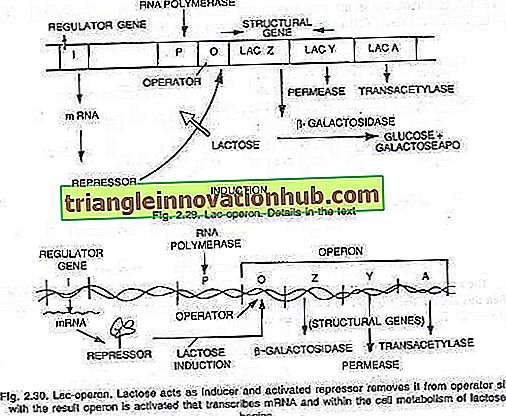
Francois Jacob e Jacques Monod (1961), mentre lavoravano all'Istituto Pasteur di Parigi, proposero che le vie metaboliche fossero regolate come unità. Hanno studiato il cambiamento nella natura degli enzimi di E. coli se forniti con diversi tipi di zuccheri. Hanno trovato, quando lo zucchero lattosio è aggiunto alle colture di E. coli, induce tre enzimi necessari per scomporre il lattosio in glucosio e galattosio.
Questi enzimi recentemente sintetizzati sono- (i) p-galattosidasi, (ii) permeasi e (iii) transacetilasi. Secondo loro, la sintesi di questi tre enzimi è controllata da un lungo segmento di DNA noto come operone che è divisibile in un sito dell'operatore O e tre geni strutturali Z, Y e A. L'azione dei geni strutturali è regolata dal sito dell'operatore con il aiuto di una proteina repressore prodotta dall'azione del gene 'i' noto come gene regolatore.
I geni sono espressi o non espressi che dipende dal fatto che l'interruttore dell'operatore sia acceso o spento. Quando l'interruttore è attivo, i tre geni vengono trascritti da RNA polimerasi in un unico tratto di RNA messaggero che copre tutti e tre i geni. Ogni gene strutturale è generalmente noto come cistron e il lungo mRNA trascritto che copre tutti i cistroni è noto come policistronico.
L'accensione o lo spegnimento dell'interruttore dell'operatore è ottenuto da una proteina nota come repressore. Quando questa proteina si lega all'operatore (O) e la blocca, l'interruttore viene disattivato ei tre geni (Z, Y, A) non vengono espressi (vedere la figura 2.28).
Lac-operone:
Questo è un esempio di operone inducibile. Alcune molecole di lattosio vengono aggiunte nella cellula dall'azione della permeasi enzima, una piccola quantità di cui si trova anche in condizioni represse. Queste poche molecole vengono quindi convertite in una forma attiva di lattosio che si lega al repressore (prodotto genico) e, quindi, il repressore non può più legarsi all'operatore.
Quando l'operatore è privo di repressore, l'RNA polimerasi inizia la trascrizione dell'operone legando il sito del promotore. P. RNA messaggero corrispondente a tutti e tre gli enzimi viene sintetizzato e tradotto in tre enzimi: P-galattosidasi, permeasi e transacetilasi.
Con la produzione di questi tre enzimi inizia il metabolismo del lattosio. La sintesi degli enzimi viene continuata a meno che e fino a quando tutte le molecole di lattosio vengano consumate dalla cellula. Quando vengono consumate anche le ultime molecole di lattosio, legate al repressore, il repressore inattivo diventa attivo e si lega al sito dell'operatore (O) per spegnere normalmente l'operone (vedere le figure 2.29 e 2.30).
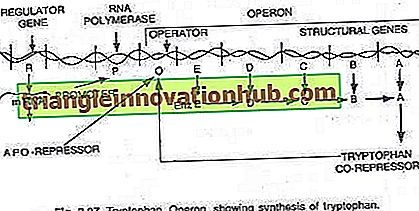
Triptofano-operone:
In questo caso, l'operone è generalmente attivo in modo che la trascrizione e la traduzione siano attive per sintetizzare gli enzimi necessari per la sintesi di un determinato metabolita dalla cellula. Tuttavia, l'operone può essere spento quando la cellula non richiede il metabolita o il metabolita è stato prodotto nell'eccesso. L'operone del triptofano è costituito da cinque geni (trp E, D, C, B e A) che codificano per cinque enzimi (En z E, D, C, B e A) catalizzando la sintesi di triptofano (un amminoacido) e quindi costituendo un percorso anabolico. La presenza del triptofano serve a reprimere la sintesi degli enzimi responsabili della sua fabbricazione (vedi figure 2.27 e 2.31).
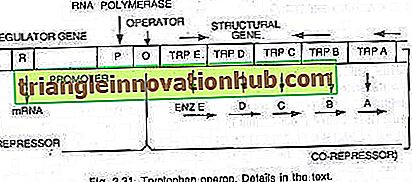
I cinque geni strutturali che codificano per questi cinque enzimi sono regolati da un singolo interruttore operatore. Qui, la proteina repressore prodotta dai geni regolatori non si lega all'operatore da sola. Tuttavia, quando viene aggiunto il triptofano, esso si lega al repressore ed è noto come co-repressore. Il complesso repressore-corepressore si lega all'operatore e blocca l'espressione di questo operone.
La struttura dell'operone del triptofano è più o meno simile a quella dell'operone lac, ma ha una variazione funzionale. In questo caso il prodotto del gene R è equivalente al gene i dell'operone lac. Il prodotto del gene R produce proteine che da sole non sono in grado di legarsi all'operatore. Questo è indicato come apo-repressore.
In presenza di triptofano un co-repressore, si forma il repressore funzionale che ora si lega all'operatore e impedisce la trascrizione dell'operone e la produzione di triptofano. Qui il modello operonico spiega sia il fenomeno di induzione e repressione nei procarioti.


