Organi linfatici e circolazione linfatica
Organi linfatici e circolazione linfatica!
Tour dei leucociti nel corpo:
I globuli bianchi (leucociti) sono prodotti dalle cellule staminali ematopoietiche nel midollo osseo negli adulti e rilasciati nella circolazione.
Dai capillari sanguigni i leucociti e il liquido nel sangue fuoriescono e penetrano negli spazi del tessuto. Il fluido negli spazi del tessuto è chiamato fluido interstiziale. Parte del liquido interstiziale ritorna alla circolazione sanguigna attraverso le vene. Un'altra parte del liquido interstiziale passa attraverso sottili vasi capillari chiamati vasi linfatici.
Il fluido nel vaso linfatico è chiamato linfa. I leucociti navigano insieme con il fluido linfatico agli organi linfoidi secondari come i linfonodi. Dai linfonodi navigano di nuovo con il fluido linfatico e rientrano nella circolazione sanguigna. Così, i leucociti pattugliano tutto il corpo viaggiando nel sangue e nella linfa. A causa della loro capacità di raggiungere quasi tutte le parti del corpo, i leucociti possono riconoscere la presenza di agenti estranei come i batteri in qualsiasi parte del corpo. Dopo il riconoscimento degli agenti estranei le risposte immunitarie sono indotte immediatamente in modo che gli agenti estranei vengano eliminati il prima possibile.
Organi linfoidi primari:
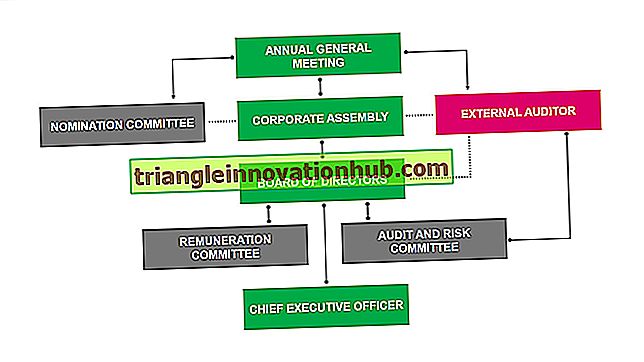
Il midollo osseo e il timo sono chiamati organi linfoidi primari o centrali (Fig. 5.1). Le cellule T e B sono prodotte dalle cellule staminali ematopoietiche pluripotenti nel midollo osseo. Tuttavia, i luoghi di maturazione delle cellule T e B sono diversi. La maturazione delle cellule B si verifica nel midollo osseo. Considerando che, la maturazione delle cellule T è completata nel timo. Ogni giorno circa 10 'linfociti maturi vengono rilasciati in circolo da timo e midollo osseo.

Fig. 5.1: organi linfoidi primari e secondari umani. Il midollo osseo e il timo sono gli organi linfoidi primari (o centrali). Milza, linfonodi e tessuti linfoidi associati alle mucose (come le tonsille, i cerotti di Peyer nell'intestino e l'appendice) sono gli organi linfoidi secondari (o periferici)
Midollo osseo:
Quasi tutte le cellule del sangue (globuli rossi, globuli bianchi e piastrine) sono derivate dalle cellule staminali ematopoietiche pluripotenti (HSC) nel midollo osseo. Il processo mediante il quale le cellule del sangue crescono, si dividono e si differenziano nel midollo osseo è chiamato ematopoiesi.
Durante la vita fetale, le CSE del fegato fetale migrano e colonizzano le cavità del midollo osseo. Per nascita, le CSE occupano praticamente l'intero spazio del midollo osseo. Dopo la nascita, il midollo osseo è il sito di produzione di cellule del sangue da parte delle CSE. Mentre l'individuo invecchia, l'attività ematopoietica nelle ossa più grandi diminuisce. Dopo la pubertà l'ematopoiesi è in gran parte confinata alle ossa scheletriche assiali (come pelvi, sterno, costole, vertebre e cranio).
Timo:
Il timo è coinvolto nella maturazione dei linfociti T. Nel timo, i linfociti T maturano e diventano funzionalmente competenti. Il timo è composto da due lobi e si trova sopra il cuore sovrastante i principali vasi sanguigni. Le cellule epiteliali del timo producono un certo numero di ormoni peptidici (come timulina, timopoietina e timosina).
Si ritiene che questi ormoni attraggono le cellule T del precursore o progenitore (rilasciate dal midollo osseo) dal sangue e successivamente le differenziano in cellule T mature. Le cellule T progenitrici interagiscono con le cellule del timo, come le cellule del timo-corticale, le cellule epiteliali del timo midollare, le cellule dendritiche interdigitanti e i macrofagi (figura 5.2).
L'interazione cellula-cellula tra lo sviluppo delle cellule T e delle cellule del timo e gli effetti degli ormoni timici portano alla maturazione delle cellule T nel timo. I linfociti T che si trovano all'interno del timo sono spesso chiamati timociti.

Fig. 5.2: Schema della sezione trasversale di una porzione di timo.
Il timo è circondato da una capsula. Molte trabecole si estendono dalla capsula al timo. La regione corticale contiene molte cellule T progenitrici, cellule infermieristiche e cellule epiteliali timiche corticali. Il midollo contiene linfociti più maturi, cellule dendritiche interdigitanti e cellule epiteliali timiche midollari. L'interazione delle cellule T progenitrici con le varie cellule nel timo e gli ormoni timici porta alla maturazione delle cellule T. Durante lo sviluppo un numero enorme di cellule T in via di sviluppo muoiono nel timo. I corpuscoli di Hassall contengono cellule epiteliali degenerative
Alcune molecole sulla superficie dei leucociti vengono utilizzate per distinguere i leucociti come cellule T, cellule B, ecc. Queste molecole sono chiamate molecole CD (cluster di differenziazione comune). (Per esempio, le molecole CD4 sono presenti sulla superficie delle cellule T helper e quindi le cellule T helper sono chiamate cellule T CD4 + . Allo stesso modo, le molecole CDS sono presenti sulla superficie delle cellule T citotossiche e quindi, le cellule T citotossiche sono dette cellule CD8 + T).
I linfociti T precoci rilasciati dal midollo osseo entrano nel timo. Le cellule T premature rilasciate dal midollo osseo non esprimono molecole di superficie CD4 e CDS (Figura 5.3) e quindi, sono chiamate cementi double negative (CD4 CD8 - ovvero entrambe le molecole CD4 e CD8 non sono presenti sulle loro membrane cellulari).
Durante il loro sviluppo iniziale entrambe le molecole CD4 e CDS appaiono sulla loro membrana cellulare (e quindi ora si dice che siano cellule a doppia positività; CD4 + CD8 + ).
Durante l'ulteriore sviluppo, ciascuna doppia cellula positiva interrompe selettivamente l'espressione della molecola CD4 o CDS. Di conseguenza, esprimono una qualsiasi delle molecole sulla loro superficie e quindi le cellule ora diventano singole cellule positive, (CD4 + CD8 - o CD4 - CD8 + ).
Le cellule, che diventano singole positive, lasciano il timo come linfociti T maturi nella circolazione sanguigna.
I meccanismi esatti alla base di questi eventi non sono chiaramente noti. Sorprendentemente, quasi il 99 percento delle cellule doppie positive muore all'interno del timo. Le cellule restanti maturano in cellule singole positive (CD4 + CD8 - o CD4 - CD8 + ) e lasciano il timo come cellule T mature. Non sono note le ragioni e i meccanismi alla base della morte di un numero enorme di cellule doppie positive nel timo. Si ritiene che le cellule T auto-reattive muoiano nel timo in modo che non si verifichi una risposta autoimmune (cioè le risposte immunitarie contro le auto-molecole).

Fig. 5.3: Sviluppo dei linfociti T nel timo.
I linfociti T sono prodotti da cellule staminali ematopoietiche nel midollo osseo. I linfociti T rilasciati dal midollo osseo in circolo non sono linfociti T maturi e sono chiamati linfociti T progenitori. I linfociti T progenitori entrano nel timo, dove lo sviluppo dei linfociti T è completato.
La cellula T progenitrice che entra nel timo non esprime molecole CD4 e CDS sulla sua superficie cellulare (e quindi chiamate cellule doppie-negative; CD4 - CD8 - ). Mentre la cellula si sviluppa, entrambe le molecole di CD4 e CDS appaiono sulla sua superficie (e quindi la cellula è chiamata cellula doppio-positiva; CD4 + CD8 + ). Man mano che la cellula si sviluppa ulteriormente, la cellula spegne l'espressione di molecola CD4 o CD8 ed esprime una qualsiasi delle molecole sulla superficie cellulare (e quindi chiamata singola cellula positiva: CD4 + CD8 - o CD4 - CD8 + ). Le cellule T mature, singole e positive vengono rilasciate dal timo nella circolazione sanguigna
Il timo funziona anche nel terzo mese di vita fetale. Alla nascita, il timo è molto attivo. Continua ad allargarsi per molti anni e raggiunge il suo picco di peso alla pubertà. Successivamente si evolve. C'è una diminuzione nel numero di linfociti. C'è un'atrofia delle cellule epiteliali del timo e sono sostituite dal grasso. Dall'età di 40 a 45 anni, i tessuti grassi sostituiscono più del 50 per cento del timo.
La completa assenza congenita del timo determina l'assenza di linfociti T e causa una grave malattia da immunodeficienza potenzialmente letale. L'assenza di timo congenita nell'uomo causa la sindrome di DiGeorge. I topi con assenza congenita di timo sono chiamati topi nudi.
Organi linfoidi secondari:
I linfociti B maturi rilasciati dal midollo osseo e i linfociti T maturi rilasciati dal timo sono in uno stato "quiescente" o "a riposo" e sono chiamati linfociti "vergini" o "ingenui". I linfociti vergini migrano in vari organi linfoidi secondari (o periferici) come milza, linfonodi o tessuti linfoidi associati alla mucosa (MALT).
Gli organi linfoidi secondari aiutano a contatto tra linfociti e sostanze estranee, portando all'attivazione dei linfociti contro sostanze estranee. Dopo l'attivazione, i linfociti subiscono la divisione cellulare e svolgono molte funzioni immunologiche.
I materiali estranei sono solitamente chiamati antigeni. Gli organi linfoidi secondari sono strettamente stipati con linfociti e cellule presentanti l'antigene (macrofagi e cellule dendritiche).
io. L'imballaggio stretto delle cellule immunitarie negli organi linfoidi secondari aiuta a trattenere il materiale estraneo negli organi linfoidi secondari.
ii. L'imballaggio stretto delle cellule immunitarie aiuta anche il contatto dell'antigene con le cellule immunitarie e la conseguente attivazione delle cellule contro gli antigeni. (Ad esempio, i batteri che entrano attraverso una lesione del dito vengono trasportati insieme al liquido linfatico ai linfonodi locali dell'ascella mentre passano attraverso i linfonodi, i batteri sono trattenuti nei linfonodi. i siti, in cui le cellule immunitarie sono strettamente legate, portano al contatto dei batteri con le cellule immunitarie, che provocano l'attivazione dei linfociti e il successivo sviluppo di risposte immunitarie contro i batteri).
La maggior parte delle risposte immunitarie contro sostanze estranee vengono lanciate dagli organi linfoidi secondari.
Linfonodi:
Dai capillari sanguigni, i leucociti e il liquido del sangue fuoriescono negli spazi dei tessuti. Il fluido negli spazi del tessuto è chiamato fluido tissutale interstiziale. Una parte del fluido interstiziale passa attraverso sottili vasi capillari chiamati vasi linfatici e il fluido all'interno dei vasi linfatici viene chiamato linfa.
Durante il suo passaggio, la linfa scorre attraverso una serie di piccoli organi a forma di fagiolo chiamati linfonodi, che sono distribuiti lungo l'intera lunghezza dei vasi linfatici. Spesso si presentano come catene o grappoli e ricevono linfa da un particolare organo o regione del corpo.
Il linfonodo funziona come un filtro fisico e biologico. Linfonodo è riempito con densi aggregati di linfociti, cellule dendritiche e macrofagi. La linfa entra nel nodo attraverso molti vasi linfatici afferenti (in arrivo), percola attraverso le cellule impaccate ed esce attraverso il vaso linfatico efferente (uscente) sul lato opposto (Fig. 5.4) del linfonodo.
Il linfonodo ha tre regioni chiamate corteccia, paracortex e midollo (Fig. 5.4).
Nella corteccia, ci sono diverse aree discrete sferiche o ovoidali chiamate follicoli linfoidi. I follicoli linfoidi sono composti principalmente da cellule B, alcune cellule T (tutte sono cellule T helper) e speciali tipi di cellule chiamate cellule dendritiche follicolari.
Ci sono due tipi di follicoli linfoidi chiamati follicoli linfoidi primari e follicoli linfoidi secondari. Prima della stimolazione antigenica, le cellule B nel follicolo linfoide sono in uno stato di riposo e il follicolo linfoide è chiamato follicolo linfoide primario. Gli antigeni (come i batteri) che entrano attraverso la pelle o la membrana mucosa vengono trasportati insieme alla linfa ed entrano nel linfonodo. Le cellule B del follicolo primario si legano agli antigeni trasportati dalla linfa. Il legame degli antigeni con le cellule B avvia l'attivazione delle cellule B.
Dopo l'attivazione delle cellule B, il follicolo primario è chiamato follicolo linfoide secondario. Le cellule B attivate nel follicolo secondario si dividono rapidamente e viene prodotto un numero enorme di cellule. L'area centrale del follicolo linfoide secondario contiene cellule B che si dividono rapidamente e quest'area è chiamata centro germinativo. Il centro germinale contiene linfociti, la maggior parte dei quali si trovano in diversi stadi di attivazione e trasformazione dell'esplosione. L'area periferica o del mantello contiene cellule B mature.

Fig. 5.4: Diagramma della sezione trasversale del nodo linfatico che mostra i follicoli del lymplioid primario e secondario.
Il linfonodo è circondato da una capsula. Molti vasi linfatici afferenti (che drenano la linfa dagli spazi dei tessuti) entrano nel linfonodo. Il fluido linfatico e gli antigeni (se presenti) dai tessuti entrano nel linfonodo attraverso i vasi linfatici afferenti. La linfa e gli antigeni percolano attraverso le cellule strettamente impacchettate nel linfonodo. Il linfonodo ha la corteccia, la corteccia para e le regioni midollari. Nella corteccia sono presenti follicoli linfoidi primari (costituiti da molte cellule B di riposo) e follicoli linfoidi secondari (costituiti principalmente da cellule B attivate in rapida divisione).
L'area della corteccia Para contiene cellule T, macrofagi e cellule dendritiche interdigitanti. L'area midollare più interna contiene poche cellule linfoidi. Durante il passaggio della linfa e degli antigeni dai vasi linfatici afferenti verso il vaso linfatico efferente gli antigeni vengono filtrati e raccolti dai macrofagi / cellule dendritiche / cellule B. Di conseguenza, le risposte immunitarie si sviluppano contro l'antigene.
I follicoli linfoidi secondari non sono presenti alla nascita perché il feto nella madre non è solitamente esposto a batteri o virus. Dopo la nascita, i follicoli linfoidi secondari si sviluppano a causa dell'esposizione ripetuta a sostanze estranee come i batteri. La presenza di un follicolo secondario in un linfonodo rappresenta una risposta immunitaria in corso.
Le cellule B attivate nel follicolo linfoide secondario si dividono ripetutamente per produrre plasmacellule e cellule B di memoria. Le plasmacellule nei follicoli secondari secernono anticorpi e gli anticorpi sono trasportati insieme al flusso linfatico nel flusso sanguigno. Le cellule follicolari dendritiche nel follicolo linfoide sono responsabili dell'assemblaggio delle cellule della memoria nei follicoli linfoidi e regolano le loro attività successive.
L'area del linfonodo paracellulare contiene cellule T, macrofagi e cellule dendritiche interdigitanti. Le cellule e i macrofagi interdigitanti intrappolano gli antigeni nella linfa e presentano gli antigeni alle cellule T helper. Di conseguenza, le cellule T helper vengono attivate e le cellule T helper attivate innalzano le risposte immunitarie contro gli antigeni.
Il midollo è lo strato più interno del linfonodo e quest'area contiene poche cellule linfoidi, principalmente plasmacellule.
I vasi linfatici dai tessuti (chiamati vasi linfatici afferenti) drenano la linfa nella corteccia del linfonodo. La linfa percola attraverso la corteccia e le aree della corteccia di Para e fluisce fuori dal linfonodo attraverso il vaso linfatico efferente. Durante il flusso di linfa dalla corteccia al vaso linfatico efferente, la linfa percolata attraverso le cellule immunitarie, e questo aiuta a filtrare gli antigeni e il successivo contatto tra antigeni e cellule immunitarie (come cellule B, cellule dendritiche follicolari e cellule T ).
I linfociti e le cellule dendritiche follicolari (che fungono da cellule antigene-presentanti) sono strettamente stipati nei linfonodi. Questo microambiente aiuta nella comunicazione efficace (tramite citochine e contatto cellula-cellula) tra queste cellule, portando all'induzione delle risposte immunitarie contro gli antigeni trattenuti nel linfonodo. Pertanto, molte delle risposte immunitarie contro gli antigeni estranei sono indotte nei linfonodi.
Milza:
La milza si trova appena sotto il diaframma sul lato sinistro dell'addome e pesa circa 150 g nell'adulto. Quando il sangue passa attraverso la milza, la milza filtra e intrappola nel sangue gli antigeni estranei (come i microbi). Pertanto, la milza svolge un ruolo importante nel controllare la diffusione dei microbi in diverse parti del corpo attraverso il sangue.
La milza è circondata da una capsula. La capsula della milza estende un numero di sporgenze all'interno della milza per formare un certo numero di compartimenti. Ci sono due tipi di compartimenti nella milza chiamati polpa rossa e polpa bianca.
io. Nell'area della polpa rossa, i vecchi globuli rossi e i globuli rossi difettosi vengono distrutti.
ii. Molte cellule T, cellule B e cellule dendritiche interdigitanti popolano l'area della polpa bianca. Le cellule B sono organizzate in follicoli linfoidi primari. Dopo la sfida antigenica, i follicoli primari si sviluppano in follicoli linfoidi secondari. Le cellule dendritiche interdigitanti della milza intrappolano gli antigeni nel sangue e li presentano alle cellule T helper, portando all'attivazione delle cellule T helper. Le cellule T helper attivate aiutano nell'attivazione delle cellule B.
Tessuti linfoidi associati alla mucosa:
Le vie respiratorie, il tratto gastrointestinale e il tratto genito-urinario sono coperti da membrane mucose. Molti microbi possono entrare attraverso la membrana mucosa nel corpo. Pertanto, le forze di difesa sono necessarie in questo punto di ingresso vitale per contrastare i microbi a livello della mucosa stessa. I tessuti linfoidi che difendono questa vasta area sono collettivamente chiamati tessuti linfoidi associati alla mucosa (MALT). Esistono due tipi di disposizione dei tessuti linfoidi nelle aree della mucosa.
1. Le cellule linfoidi sono disposte come ammassi liberi
2. I tessuti linfoidi sono organizzati come strutture organizzate (come le tonsille, l'appendice e i cerotti di Peyer).
Patch di Peyer nella mucosa intestinale:
Le cellule epiteliali della mucosa rivestono l'aspetto esterno della mucosa intestinale (Figura 5.5). Ci sono linfociti nello strato epiteliale della mucosa e sono chiamati linfociti intraepiteliali (IEL). Molti degli IEL sono cellule T CD8 + e esprimono insoliti recettori cellulari γδT. La funzione degli IEL non è nota.
La lamina propria si trova sotto lo strato epiteliale della mucosa (Figura 5.5). La lamina propria contiene un gran numero di ammassi liberi di cellule B, plasmacellule, cellule T helper attivate e macrofagi.
Sotto la lamina propria si trova lo strato sottomucoso. Lo strato sottomucoso contiene i cerotti di Peyer. La patch di Peyer è un nodulo di 30-40 follicoli linfoidi. Come i follicoli linfoidi in altri siti, i follicoli linfoidi del cerotto di Peyer si sviluppano anche in follicoli secondari, quando vengono messi alla prova con il microbo.

Fig. 5.5: Schema della sezione trasversale dell'intestino tenue.
L'intestino tenue ha quattro strati: 1. strato epiteliale mucoso, 2. lamina propria, 3. strato sottomucoso e 4. strato muscolare. Lo strato epiteliale della mucosa consiste in un singolo strato di cellule epiteliali. Le cellule epiteliali hanno numerose dita sottili come proiezioni chiamate villi sul lato del lume intestinale. Tra le cellule epiteliali si trovano cellule specializzate chiamate cellule M. Cluster sciolti di follicoli linfoidi (costituiti da un gran numero di cellule B, plasmacellule, cellule e macrofagi) sono presenti nella lamina propria. I cerotti di Peyer sono presenti nello strato sottomucoso. Il cerotto di Payer consiste di 30-40 follicoli linfoidi
Le cellule specializzate chiamate cellule M (figura 5.6) si trovano nello strato epiteliale della mucosa. Le cellule M non hanno microvilli (mentre le cellule epiteliali hanno microvilli). Le cellule M sono cellule epiteliali appiattite e hanno una invaginazione profonda o una tasca nell'aspetto basolaterale della membrana plasmatica. Questa tasca contiene cellule B, cellule T e macrofagi.
L'antigene (come i batteri) nel lume intestinale viene portato nella cellula M.
I batteri vengono quindi trasportati sull'altro lato della cellula M e rilasciati nella tasca della cellula M.
Le cellule B e le cellule T nei follicoli linfoidi vicino alle cellule M riconoscono i batteri e si attivano.
Di conseguenza, le risposte immunitarie sono indotte contro i batteri. Le cellule B attivate si differenziano in plasmacellule e secernono immunoglobulina A (IgA).
Le IgA vengono trasportate dalle cellule epiteliali della mucosa del lume intestinale dell'irito (Fig. 5.6), dove l'IgA si lega al microbo e impedisce l'ingresso del microbo attraverso la mucosa.
Tessuti linfoidi nella pelle:
Un piccolo numero di linfociti sono costantemente presenti nel derma e nell'epidermide della pelle. L'epidermide contiene anche cellule chiamate cellule di Langerhans, che funzionano come cellule presentanti l'antigene. Quando incontrano sostanze estranee, le cellule di Langerhan le inghiottono e navigano insieme al fluido linfatico verso il linfonodo locale. Le cellule di Langerhan esprimono livelli molto alti di molecole MHC di classe II e presentano l'antigene alle cellule T helper nel linfonodo.

Fig. 5.6: Cella M.
Le cellule M sono cellule epiteliali specializzate situate lungo le cellule epiteliali della mucosa dei tratti gastrointestinali, respiratori e genitourinari. Le cellule M trasportano i microbi dal lume gastrointestinale, respiratorio e genito-urinario nel corpo. La cellula M inghiotte il microbo nel lume intestinale.
Il microbo inghiottito viene trasportato attraverso la cellula M. La membrana della vescicola endocitica si fonde con la membrana delle cellule M e rilascia il microbo nella tasca della cellula M. Le cellule T, le cellule B, i macrofagi e le cellule dendritiche nei follicoli linfoidi sottostanti riconoscono il microbo. Di conseguenza, le risposte immunitarie sono indotte contro il microbo.
Le cellule B attivate nei follicoli linfoidi si dividono per produrre plasmacellule e le plasmacellule a loro volta secernono anticorpi IgA specifici contro il microbo. L'IgA viene trasportata dalle cellule epiteliali mucose nel lume.
L'IgA si lega al microbo specifico nel lume e interferisce con l'ingresso del microbo attraverso la membrana mucosa (l'endocitosi è un processo mediante il quale le cellule inglobano macromolecole extracellulari Una piccola porzione di membrana plasmatica circonda la macromolecola e racchiude la macromolecola. le membrane si fondono e si staccano per formare una vescicola contenente la macromolecola)
Circolazione linfatica:
Il sangue circola a causa della pressione creata dall'azione di pompaggio del cuore. I capillari sanguigni hanno pareti molto sottili. A causa della pressione all'interno dei capillari sanguigni, il fluido dal sangue filtra dai capillari negli spazi dei tessuti. Il fluido nel tessuto è chiamato fluido interstiziale.
Parte di questo fluido ritorna direttamente al flusso sanguigno attraverso i venule del sangue e il liquido rimanente scorre attraverso gli spazi del tessuto e si raccoglie nei canali a parete sottile chiamati vasi linfatici (Fig. 5.7).
Il fluido nei vasi linfatici è chiamato linfa. La linfa scorre lentamente e raggiunge i linfonodi. Dai linfonodi la linfa scorre ulteriormente ed entra nella circolazione sanguigna attraverso la vena succlavia sinistra nel torace. Pertanto, i vasi linfatici fungono da sistema di drenaggio che raccoglie il fluido dagli spazi del tessuto e restituisce il fluido al flusso sanguigno.

Fig. 5.7: circolazione linfatica.
Il fluido negli spazi del tessuto è chiamato fluido interstiziale. Una parte del liquido interstiziale entra in vasi capillari sottili chiamati vasi linfatici. Il fluido nei vasi linfatici è chiamato linfa. La linfa scorre lungo il vaso linfatico afferente ed entra nei linfonodi. Dai linfonodi la linfa passa attraverso i vasi linfatici efferenti. I vasi linfatici di molte parti del corpo si uniscono e formano un vaso linfatico più grande chiamato dotto toracico. Il dotto toracico drena la linfa nella circolazione sanguigna attraverso la vena succlavia sinistra.
La linfa contiene molte sostanze difensive e globuli bianchi, che esaminano l'intero corpo circolando nelle circolazioni sanguigne e linfatiche. Durante il loro tour, i globuli bianchi e altre sostanze attaccano qualsiasi intruso estraneo (come i batteri) e li rimuovono, in modo che l'uomo viva una vita più sana.
I vasi linfatici sono canali estremamente delicati attraverso i quali scorre la linfa. La linfa che passa attraverso un'area di infezione batterica porterà i batteri insieme ai linfonodi locali. Il linfonodo agisce come un filtro e blocca i batteri. Pertanto, il linfonodo previene la diffusione di batteri in altre parti del corpo.
Il linfonodo contiene numerose cellule presentanti l'antigene (APC) -T linfociti e linfociti B. Queste cellule difensive riconoscono gli antigeni batterici e innalzano le risposte immunitarie contro i batteri, che portano alla distruzione dei batteri.
Se i batteri escono dal linfonodo, i batteri entrano nella circolazione sanguigna e possono raggiungere qualsiasi parte del corpo. In tali situazioni, i macrofagi nel fegato e nella milza svolgono un ruolo importante nel catturare i batteri e prevenire la diffusione di batteri nel sangue.
I linfociti sono cellule migratorie, ovvero si spostano da un luogo all'altro. Ad esempio, un linfocita individuale può rimanere in un linfonodo per 12 ore. Quindi si stacca dal linfonodo ed entra nella circolazione sanguigna, dove rimane per pochi minuti o poche ore. Dalla circolazione sanguigna, si spostano su qualsiasi altro tessuto o linfonodo. Con la sua capacità di viaggiare in qualsiasi parte del corpo, i linfociti esaminano l'intero corpo, durante il giorno e la notte, alla ricerca di sostanze estranee. (Come la polizia che viaggia in ogni angolo della città in cerca di ladri che potrebbero essere entrati in città.)
Se i linfociti negli organi linfoidi secondari incontrano sostanze estranee, i linfociti vengono attivati contro la particolare sostanza estranea. I linfociti attivati si dividono per produrre molte cellule figlie. Alcune delle cellule figlie diventano cellule effettrici e altre diventano celle di memoria.
Le cellule effettrici sono di breve durata e funzionano per la rimozione immediata di antigeni estranei. Mentre le celle di memoria hanno molti anni di vita e funzionano durante il successivo ingresso della stessa sostanza estranea nel corpo (in modo che la sostanza estranea venga rimossa prima che possa produrre alcun danno).
Le cellule di memoria e le cellule effettrici hanno una forte preferenza a ritornare allo stesso tipo di tessuto in cui si è verificata la loro attivazione. Ad esempio, una cellula di memoria sviluppata nell'intestino (in risposta ai batteri che entrano attraverso l'intestino) avrà la tendenza a migrare verso il tessuto linfoide associato all'intestino per il resto della sua vita, che potrebbe durare molti anni. Restando in questa zona, danno protezione attivandosi ogni volta che i particolari batteri entrano attraverso l'intestino.
Rilevanza clinica:
Infezione batterica acuta e linfoadenite:
Durante le infezioni batteriche acute della pelle e dei tessuti sottocutanei i microbi vengono trasportati lungo la linfa ai linfonodi locali. Di conseguenza, i linfociti nei linfonodi locali vengono attivati e si instaura una reazione infiammatoria. Vi è un aumento del flusso sanguigno, il rilascio di mediatori dell'infiammazione e la cessazione della normale emigrazione dei linfociti dal linfonodo, con conseguente ingrossamento dei linfonodi. I linfonodi infiammati sono grandi, dolorosi e teneri e indicati come linfoadenite. In generale, la presenza di linfonodi ingrossati, dolorosi e dolorosi suggerisce un'infezione batterica acuta.
Splenectomia e batteriemia:
Come parte del trattamento per alcune malattie, la milza dei bambini viene rimossa dall'intervento chirurgico (chiamato splenectomia). Nei bambini splenectomizzati, vi è un'aumentata incidenza di alcune malattie batteriche (causate da Streptococcus pneumoniae, Neisseria meningitidis e Haemophilus influenzae).
Questi batteri causano infezioni gravi e possono diffondersi attraverso il sangue. A causa dell'assenza di milza nei bambini splenectomizzati, la diffusione di batteri attraverso il sangue non viene prevenuta e quindi, le probabilità di batteriemia dovuta a questi batteri sono maggiori. Di conseguenza, i bambini splenectomizzati soffrono di queste infezioni batteriche.